|
В пасти - закон эволюции
Какова же она, система развития акул? Об этом долго спорили
учёные, было придумано множество интересных гипотез и псевдонаучных
штучек. Одна из самых любопытных систем (причём, с точки зрения
многих, очень разумная) - система С. Мосса, основанная на строении
пасти.
Основываясь на строении ротового аппарата, черепа и способов
питания множества акул - как древних, так и современных - учёный
разработал такую систему. От исходных хватающих рыб произошли сосуще-хватающие
скаты (новая идея!). От них - три ветви. Ветвь А дала катранообразных,
пилоносов и морских ангелов (режущий тип пасти). Ветвь Б - сосуще-давящие
скаты-хвостоколы, от которых идут сосуще-давящие ковровые акулы
и фильтраторы (китовая, гигантская, большеротая акулы, некоторые
скаты). Ветвь В дала колющих кархариновых и ламноидных акул, которые
опять же являются предками сосуще-давящих ковровых акул. Другие
же потомки колющих - дробящие куньи акулы, с одной стороны, и опять
же фильтраторы, с другой. То есть С. Мосс разбивает вдребезги понятия
"акула" и "скат" и предлагает устанавливать
запутанные "мостики" между группами. Но самое удивительное
в том, что эта система в целом согласуется с системой, основанной
на строении головного мозга! Так что, несмотря на все "но",
оставлять гипотезы С. Мосса в стороне нельзя.
Хвост - всему голова
Раз уж я рассказал о любопытной системе С. Мосса, упомяну
о ещё одной гипотезе - гораздо более "старой", в свете
новейших находок невероятной, но всё же очень интересной, - гипотезе,
изложенной в книге В. Гааке. Он считает предками всех рыб, а заодно
и наземных позвоночных, скатов. Прежде всего, он считает, что из
продольных плавников по бокам тела развились плавники прочих рыб
и конечности наземных существ. Далее, в пользу гипотезы о появлении
наземных позвоночных из скатов Гааке утверждает, что скаты из-за
формы и способа "залегания" на дне могли оставаться после
отлива на берегу в лужицах и, таким образом, приспосабливаться к
жизни на суше! Однако главным доводом учёного является форма хвоста
скатов: хвост. Гааке утверждает, что длинный хвост является признаком
низкой ступени развития! Интересно, почему он не вспомнил о высокоразвитой
акуле-лисице (к которой мы ещё вернёмся), своим длиннющим хвостом
глушащей рыбу?
Из всех этих доводов Гааке делает "простой и ясный"
вывод: скатоподобные сначала вышли на сушу, а затем дали четыре
ветви предков: одна повела к панцирным, вернувшимся в воду, другая
- к предкам современных селахий, третья - к двоякодышащим, "наполовину
вернувшимся в воду", а четвёртая - к земноводным! А хвост скатов,
по мнению ислледователя, загнулся во время ползания сухопытных скатов
вверх - и потому хвостовые плавники у всех рыб, их "потомков",
также загибается кверху!
Гипотеза Гааке, конечно, устарела и не выдерживает критики,
однако она всё же любопытна, и обойти её вниманием, тем более что
она связана с хрящевыми рыбами, я не мог.
Или мозг?
А теперь вкратце расскажу о том, как эволюционировал мозг
акул. Для этого изучим реконструкцию мозга ископаемой акулы, мозг
синей акулы и атлантической кошачьей.
Что заметно прежде всего? Конечно, у ископаемой акулы меньше,
чем у любой из современных, обонятельные луковицы, мозжечок. Значит,
она не обладала нюхом синей или кошачьей акулы и хуже, чем они,
ориентировалась в толще воды. Слабее развит и продолговатый мозг.
Осмотрев всё внимательно, сделаем такие выводы:
1) В зависимости от степени развития акулы мозжечок разбит
на большее число долей.
2) Некоторые признаки - например, громадные обонятельные луковицы,
- можно отнести на счёт её "специальности" как приспособление,
но древней ХИЩНОЙ акуле всё же требовались развитые обонятельные
луковицы; значит, можно использовать в целях исследования величину
обонятельных луковиц.
3) Чем более развитой является акула, тем больше объём всего
мозга, со всеми его отделами.
Но стоп! Слишком увлекаясь пастями, хвостами и мозгами акул,
мы забыли узнать, какие же акулы обитали в водоёмах девонского периода.
Исправляюсь!
Одна из древнейших акул
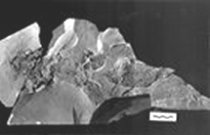 |
| Копия фрагмента скелета кладоселахии
Cladoselache fyleri |
В богатых среднедевонских отложениях штата
Огайо (США) найдены останки одних из самых древних известных акул.
Самые интересные из этих останков принадлежат кладоселахии - акуле,
составляющей особую группу, которая, по системе американского палеонтолога
Б. Шеффера, основанной на строении головного мозга, произошла напрямую
от плакодерм и не оставила известных нам потомков. Жила она отнюдь
не недавно - 375 млн лет назад. Среди находок останков кладоселахий
особенно выделяется отпечаток её тела, на котором, помимо скелета,
можно различить мышцы и почки рыбы! Отпечаток этот был найден в
Кливлендских сланцевых холмах, которые, видимо, когда-то представляли
собой дно водоёма. Помимо этого, были найдены также зубы этих акул.
Удалось установить, что кладоселахии были некрупны - 0,45-1,2 м,
изредка, вероятно, до 2 м в длину - и обладали торпедовидным обтекаемым
телом, позволявшим быстро передвигаться в поисках добычи. На спине
у кладоселахий было два плавника с острыми шипами, неравнолопастный
хвост был похож на хвост современной акулы мако и усиливался в верхней
лопасти продолжением спинного столба. Впрочем, ширина грудных плавников
говорит о том, что кладоселахии были медлительнее современных сородичей.
Глаза были большими, круглыми и, видимо, неплохо видели. Они были
настоящей грозой рыб того времени! Охотиться стаями кладоселахиям
было незачем, так как одна подобная ей рыбина могла запросто уничтожить
стайку более мелких морских обитателей (эта акула была морским жителем).
О рационе кладоселахий говорят остатки рыбы в желудках найденных
экземпляров. Но более крупные обитатели были ей страшными врагами
- например, плакодермы-артродиры вроде шестиметрового динихтиса
и девятиметрового дунклеостеуса. На зубах кладоселахий заметны следы
износа - они, конечно, не сменялись так часто, как это происходит
у современных акул.
В некоторых системах, например, созданной в 1940 г. Л. Бергом,
кладоселахии, как и ктеноканты, выделяются в особые подклассы хрящевых;
но такие взгляды, конечно же, устарели.
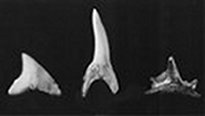 |
| Зубы кладодуса (справа) и
современных кархариновых акул |
Загадочные зубы кладодусов
Другая необычайно интересная находка, относящаяся к
среднему девону Огайо, - зубы загадочной акулы кладодуса. Зубов
было найдено лишь несколько; у каждого было по одному острию. Необыкновенная
острота этих зубов (возможно, что они принадлежали одной особи)
позволяет предположить, что кладодусы были опасными врагами мелкой
рыбы. Кладодусов близки к ктенокантам; а поскольку ктеноканты считаются
потомками или, по крайней мере, "братьями" акул-ксенакантов
(см. чуть ниже), возможно, что уже в
среднем девоне существовали и ксенаканты. Интересно, что некоторые
исследователи - например, доктор Самуэль Гарман, впервые описавший
плащеносных акул, - считали, что эти самые плащеносные акулы являются
чуть ли не современными кладодусами. Как показали более поздние
исследования, это о-о-очень далеко от истины…
Ктенокант
Близкие родственники кладодусов и о-очень дальние - кладоселахий,
ктенокантовые акулы были одними из самых распространённых пресноводных
акул девонского-каменноугольного периодов. Названа вся эта группа
по известнейшему её представителю - ктеноканту. Ктеноканты жили
в речушках и прудах девона. Их тело было, как и у кладоселахий,
торпедовидно, обтекаемо, но одно новшество сразу бросалось в глаза.
На затылке у ктеноканта располагался длинный шип, которым, несомненно,
можно было пользоваться как опасным оружием при обороне от врагов.
Впрочем, врагов у ктеноканта было не так-то много. Он был крупнее
кладоселахий, до 1,5-2 м в длину, у него были более развиты для
гребли брюшные плавники. Немного, наверное, было рыб, которые отваживались
противостоять ктеноканту! У ктенокантов, видимо, не было плакоидной
чешуи, а на родство их с хрящевыми рыбами показывают останки птеригоподий,
или гоноподий - акульих половых органов.
 |
| Ксенакант |
Откуда пошли шипы у ксенакантов
Немного я ошибся с порядком описания. Всё-таки ктеноканты - потомки
ксенакантов. Но и ксенаканты уже известны из девона. Размером примерно
с ктеноканта, пресноводная акула ксенакант также отличалась мощным
затылочным шипом. Охотились ксенаканты на мелких рыбёшек-акантодий,
стайки которых сновали в пресных водоёмах девона, и на рыб покрупнее
- плакодерм из группы антиарх и костных рыб, вроде двоякодышащих
диптеров. По моему личному мнению, ксенаканты - потомки плакодерм,
у которых были только головной и грудной костные шиты. Постепенно
костный панцирь этих плакодерм стал редуцироваться, пока не остался
только затылочный щиток, преобразовавшихся в шип. Кладоселахии же
произошли от тех же предков, что и предки гибодонтов и большинства
современных отрядов акул (о них чуть ниже). Хотя предки ксенакантов
и гибодонтов+современных акул, несомненно, близки друг другу. Ещё
в 20-е гг. XX в. А. Северцов, подметив это, предложил объединить
плевракантодов (так тогда называли ктенокантов), кладоселахий и
современных акул (селахий) в одну группу.
Модная причёска стетакантуса
Да, да, я не спорю - и кладоселахии, и ктеноканты, и ксенаканты
весьма необычны. Но они ничего не стоят по сравнению с одной из
удивительнейших древних акул - стетакантусом. Стетакантусы жили
на Земле более 300 миллионов лет назад. Они были невелики, примерно
с метр длиной. Голову их украшала, несомненно, очень модная для
того времени причёска из множества направленных назад шипов. Вдобавок
на спине стетакантуса находилась "щётка" - тоже из шипов,
только направленных вперёд. Вероятно, образ жизни стетакантусов
аналогичен таковому у прилипал - эти акулы, задирая голову, прикреплялись
к более крупным рыбам с помощью своих шипов. Помимо этого, у стетакантусов
были ещё два усеянных шипами выроста, которые отпугивали врагов,
поскольку напоминали какую-то пасть.
 |
| Останки стетакантуса |
 |
| Отпечаток ишнакантуса |
И прочие, прочие…
Казалось бы, я уже перечислил столько девонских акул! И всё-таки
многие разновидности остались "за кадром". И ишнакантус
с колючками на обеих спинных плавниках, оставивший после себя неплохой
отпечаток; и томиобатис, малоизученная акула, "брат" кладодуса
(если ксенакант - "мама")… Но пора от рассмотрения отдельных
видов перейти к более общим вопросам эволюции древних акул.
Моря и реки
Мы говорили здесь, что в девоне акулы населяли как пресные
водоёмы, так и моря, и океаны. Так где же они первоначально появились?
По-моему, на этом вопросе стоит остановиться!
В одном учебнике говорится, что акулы изначально появились
в морях. Этот взгляд устарел и основывается только на том, что кладоселахия
была морской жительницей. Появились все первые группы акул в речушках,
прудах, озёрах от пресноводных же плакодермов, и лишь потом, с образованием
свободных "дыр" в системе океанической фауны, перешли
в солёную воду.
Как же именно расселялись акулы в Мировом Океане? Н. Мягков
провёл исследования, основанные на строении головного мозга 60 современных
и 2 вымерших акул, и выяснили, что расселение шло примерно таким
путём, как вы сейчас прочтёте. (Его выводы были подтверждены работами
нескольких зарубежных исследователей.) Возле каждой экологической
группировки акул я указываю данные об их мозге. Каким образом? Первая
цифра, в %, - индекс переднего мозга, вторая - отношение индекса
переднего мозга к индексу продолговатого мозга, третья - отношение
индекса обонятельных луковиц к индексу зрительных долей; а ещё я
пишу о наличии борозд на теле мозжечка. (Разумеется, все данные
о мозге кладоселахий приблизительны.) Также замечу, что все-все-все
акулы из группировок, в названиях которых нет слов "донная",
"придонная" или "пилонос", держатся не у дна.
Итак:
примитивные исходные акулы вроде кладоселахий, глубина - около 350
м; менее 20 %, ок. 0,55, ок. 0,7, борозд нет.
древняя донная группировка, глубина - 10-50 м; 25 %,
ок. 1, 0,8, борозд нет.
древняя придонная группировка, глубина - 350-450 м;
ок. 20 %, 0,3, 0,7, борозд нет.
древняя глубоководная группировка, глубина - 450-1200
м; менее 20 %, 0,55, 0,85, борозд нет.
морские ангелы и пилоносы (?), глубина - 400-500 м;
25-30 %, ок. 1, ?, борозды могут быть, могут не быть.
придонно-пелагическая группировка (?), глубина - 300-1200
м; 20-30 %, 0,6-1,1, 0,3-1,0, борозд нет.
молодая глубоководная группировка, глубина - 300-1200
м; ок. 20 %, ок. 0,5, 0,3-0,8, борозд нет.
уклонившиеся от основного "пути": полярная
и тигровая акулы; 20-30 %, 0,3, 3,0, борозд нет.
молодая придонная группировка, глубина - 350-500 м;
28-35 %, 1,3-1,6, 1,3-2,2, борозды есть.
кошачьи акулы, глубина - 50-500 м; ок. 25 %, ок.
1, ?, борозды могут быть, могут не быть.
ковровые акулы, глубина - 50-500 м; 25-38 %, 1,2-1,5,
ок. 0,5, борозды есть.
неритическая группировка, глубина - 50-300 м; у серых
акул - ок. 40 %, 1,5-2,0, 0,8-0,9, борозды есть; у акул-молот -
45-55 %, 2,5-3,0, 1,5-1,8, борозды есть.
эпипелагическая группировка, глубина - 50-300
м; более 30 %, 1,2-2,0, 0,6-0,85, борозды есть.
Видимо, в особенно больших масштабах распространение и расселение
по различным биотопам у акул, по данным Ж.-И. Кусто и Ф. Кусто,
началось в пермотриасе, около 275 млн лет назад..
Кое-что о брадиодонтах
Раз уж мы говорим об эволюции акул, надо кое-что сказать и
об их ближайших родичах. Это рыбы из подкласса цельноголовых, или
слитночерепных.
Как ясно из названия, главный отличительный признак цельноголовых
рыб - это то, что верхняя их челюсть намертво срослась с черепом.
Этим они отличаются от акул. Но есть и много похожего. (Здесь я
эти признаки перечислять не буду.) Их хрящевый скелет позволяет
относить их к классу хрящевых, но их происхождение остаётся загадкой
для исследователей.
Самые древние цельноголовые, из вымершего отряда брадиодонтов,
известны из среднего девонского периода. Тогда же жили и кладоселахии,
и кладодусы, и ксенаканты… Один из первых брадиодонтов - хондрохелис
- был ну очень примитивной рыбой. Спинной плавник начинался примерно
там же, где и грудные, только на спине, и без какого-либо разделения
"перетекал" в хвостовой. Питались хондрохелисы моллюсками.
Эти создания были лёгкой добычей для крупных кистепёрых и, что не
исключено, акул.
В каменноугольном-пермском периодах жили более "продвинутые"
брадиодонты вроде гелодуса. У него было два спинных плавника, не
соединённых с хвостовым. Форма тела - более обтекаемая. Любопытно,
что в каменноугольных пластах Подмосковья прочные округлые дробящие
зубы цельноголовых соседствуют с многовершинковыми зубами мелких
акул. Кстати, ценнейшая коллекция этих зубов была собрана из отдельных
находок, принесённых в дар учёным школьниками-коллекционерами, -
а ведь зуб брадиодонта нелегко отличить от обычного округлого камешка!
(Об одном каменноугольном брадиодонте - протопирате - в следующем
разделе.)
Начало триасового периода - закат эпохи брадиодонтов. Причин
их вымирания несколько. Главная, видимо, - это появление питающихся
моллюсками акул с дробящими зубами, которые притесняли "засевших"
в ту же экологическую нишу брадиодонтов. В отложениях возрастом
примерно 245 миллионов лет назад останков брадиодонтов уже не видно.
А в позднем триасе появляются химеры. Этих цельноголовых, видимо,
потомков брадиодонтов, можно встретить и сегодня. Все три их современных
семейства были представлены в доисторической фауне; кроме того,
учёные выделили аж тринадцать ископаемых.
|
